D121 Antigen Presentation and T/NK Cell Activation Group
Bispecific antibodies targeting solid tumor antigens
Although T cell-recruiting CD3-binding bispecific monoclonal antibodies (BiMAb) are clinically effective for hematologic malignancies, the success of BiMAb targeting solid tumor-associated antigens (TAA) in carcinomas remains poor. We reason that co-stimulatory BiMAb in combination with anti-CD3 BiMAb would boost T cell responses, facilitating the targeting of weakly or heterogeneously expressed tumor antigens. We have analyzed anti-TAAanti-CD3/anti-CD28 BiMAb, recognizing breast cancer antigens including HER2, EGFR, CEA, and EpCAM. Moreover, bifunctional fusion proteins with tumor necrosis factor ligand (TNFL) superfamily members (4-1BBL, OX40L, CD70, TL1A) were tested. The functional activity of our bispecific antibodies was evaluated in cell-based assays. The anti-tumor activity was confirmed in a patient-derived 3D spheroid model using pleural effusions of breast cancer patients. Only in the presence of tumor cells, anti-CD3 BiMAb activated T cells and induced cytotoxicity in vitro, indicating strict dependence on cross-linking. Combination with anti-CD28 or TNFL bispecifics enhanced T cell proliferation, activation marker expression, cytokine secretion and tumor cytotoxicity, thereby reducing the minimally required dose to achieve T cell activation. Effective co-stimulation could be achieved by targeting a second breast cancer antigen, or by targeting fibroblast activation protein (FAP) expressed on another target cell. Immune checkpoint inhibitors further augmented BiMAb mediated co-stimulation. In patient-derived spheroids, co-stimulatory BiMAb were essential for activation of tumor-infiltrating lymphocytes and cytotoxic anti-tumor responses against breast cancer cells. Combining stimulatory and co-stimulatory BiMAb may have promising clinical application in the treatment of solid tumors.
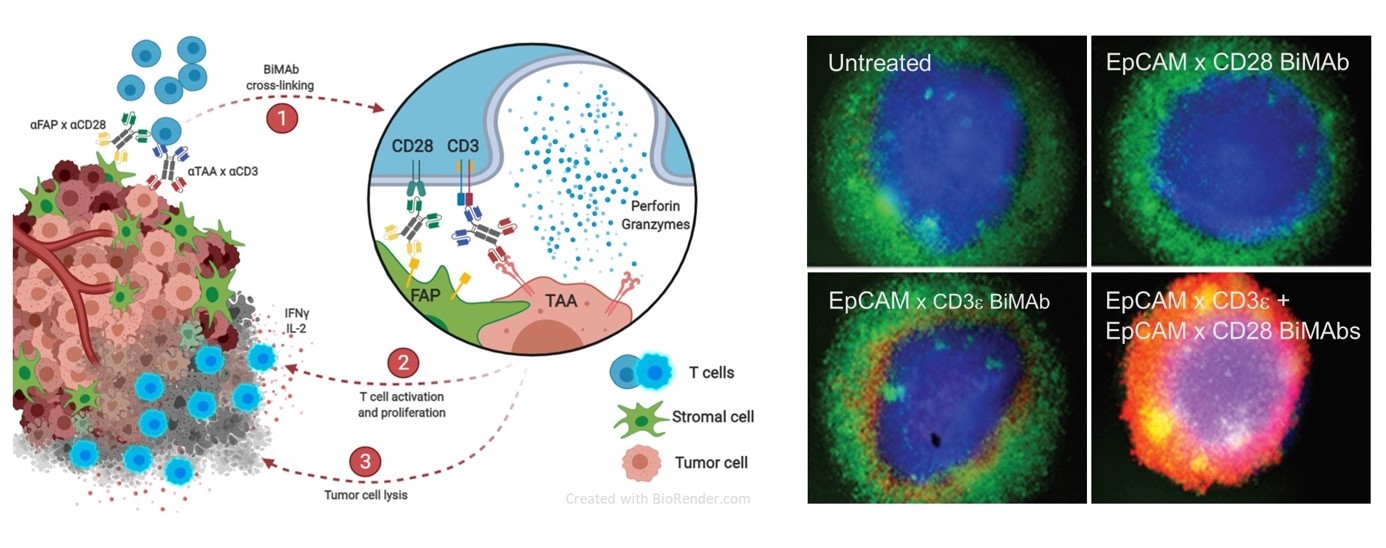
Bispecific antibodies mediate killing of breast cancer cells by T cells. Left: Graphical presentation of stimulation and co-stimulation of T cells by bispecific antibodies (BiMAb). In the split co-stimulation approach the co-stimulatory anti-CD28 bispecific antibody recognizes a tumor cell surface antigen that is different from the antigen targeted by the stimulatory anti-CD3 bispecific antibody. The former antigen can also be expressed by an adjacent stromal cell. Right: Strong tumor cell cytotoxicity is elicited by co-stimulation. Breast cancer cell organoids (blue) are cocultured with T cells (green) in the presence of BiMAb. If stimulatory and co-stimulatory BiMAb are simultaneously added, T cells efficiently invade the organoid and kill tumor cells (lower right, cell death indicated by red fluorescent dye). From: Warwas et al., Front. Immunol. 12:719116 (2021).
© DKFZ
We investigate bispecific antibodies that redirect T cells to the tumor neo-vasculature. Using VEGFR2, TIE2 and PD-L1 as target antigens expressed by proliferating endothelial cells (EC), we found that bispecific antibodies addressing either CD3 or CD28 on T cells are effective in recruiting T cells towards EC leading to a profound on-site T cell activation as well as upregulation of adhesion molecules of resting EC. This results in an augmented transendothelial migration in vitro and subsequent tumor cell killing.
Together with partners in Israel, we engineer and test novel bispecific antibodies that target NK cells towards cancer cells. Such antibodies trigger NK cells through the activating NKp46 receptor after cross-linking with, e.g., HER-2+ tumor cells and elicit tumor antigen-specific NK cytotoxicity against MHC-I-positive tumor targets. Bispecific antibodies that mediate NK cell immune checkpoint inhibition at the tumor site are in development.
We have engineered tetravalent IgG-like TCR-antibody fusion proteins that are able to redirect NK cells towards tumor cells expressing particular peptide-MHC-I complexes recognized by respective TCRs. NK cells are recruited and activated through single-chain antibodies (scFv) binding NKp46 or CD16. Furthermore, TCR-anti-CD3 scFv fusion proteins are successfully employed to elicit T cell-mediated cytotoxicity against pMHC-I-presenting tumor cells. Multimeric formats are under development in order to increase the avidity of TCRHLA-I interactions.
Recombinant fusion proteins of MHC class I molecules loaded with viral peptides and single-chain antibodies recognizing tumor-associated antigens such as EGFR or HER-2 ("HistoMAb") enable the recruitment to solid tumors of virus-peptide specific T cells with specificity for the particular peptide MHC-I complex. After showing in vitro proof-of-principle for HistoMAb with human human tumor antigen reactivity, we are in the process of establishing mouse tumor models to examine the in vivo efficacy of murine surrogate HistoMAb and xenograft models together with adoptive human T cell transfer. Furthermore, HistoMAb molecules with specificity for B-cell, T-cell and myeloid leukemia antigens will be systematically investigated.
Neoepitope-specific TCR discovery using recombinant MHC molecules
Individual mutations in tumor-expressed proteins represent attractive targets for peptide-specific T cell receptors (TCR) employed in adoptive T cell therapy regimens since they are unique to the transformed cell population. If carefully checked for absence of cross-reactivity to wild type peptides, such neoepitope-specific TCR bear a low risk of inadvertent reactivity with normal tissues, whereas TCR directed against non-mutated oncofetal or differentiation antigens can potentially pose a significant safety risk.
An effective workflow leading to the identification of neoepitope-specific T cell receptors with the goal to produce autologous TCR-transgenic T-cell products for adoptive cell therapy has been established. Using exome and tumor mRNA sequencing data, point mutations as well as frameshift mutations are identified. In silico prediction algorithms are employed to identify the most promising neoepitope ligands for each of the patients' six MHC-I (HLA-A, -B, or -C) molecules. Available immunoproteome data identifying in vivo processed and presented peptides are taken into account. We identify rare neoepitope-reactive T cells directly from peripheral blood, or after in-vitro peptide restimulation, in a highly sensitive, multiplex dual-color pMHC-I multimer staining approach followed by T cell single-cell sequencing after systematically evaluating libraries of predicted mutant peptides for successful MHC-I complex binding.
Here, we take advantage of commercially available peptide receptive MHC-I multimer platforms covering the most common HLA-A,-B,-C allelic variants as well as develop robust own strategies for an in-house production of recombinant MHC-I multimer reagents to cover all HLA-A,-B,-C allotypes and to maximize the detection of potential T cell populations. This includes the usage of MHC-I single-chain trimers tethered with mutant or wild-type peptides and more recently, we developed a peptide-loadable MHC class I platform based on soluble disulfide-stabilized beta2-microglobulin/heavy chain ectodomain single-chain dimers (ds-SCD) that can be produced in large quantities in eukaryotic cells and tailored to the individual patients' HLA allotypes with only little hands-on time. By rigorously employing peptide-loaded MHC-I multimer libraries we hope to improve success rates for the discovery of personalized neoepitope/MHC-I-specific TCR in the future.
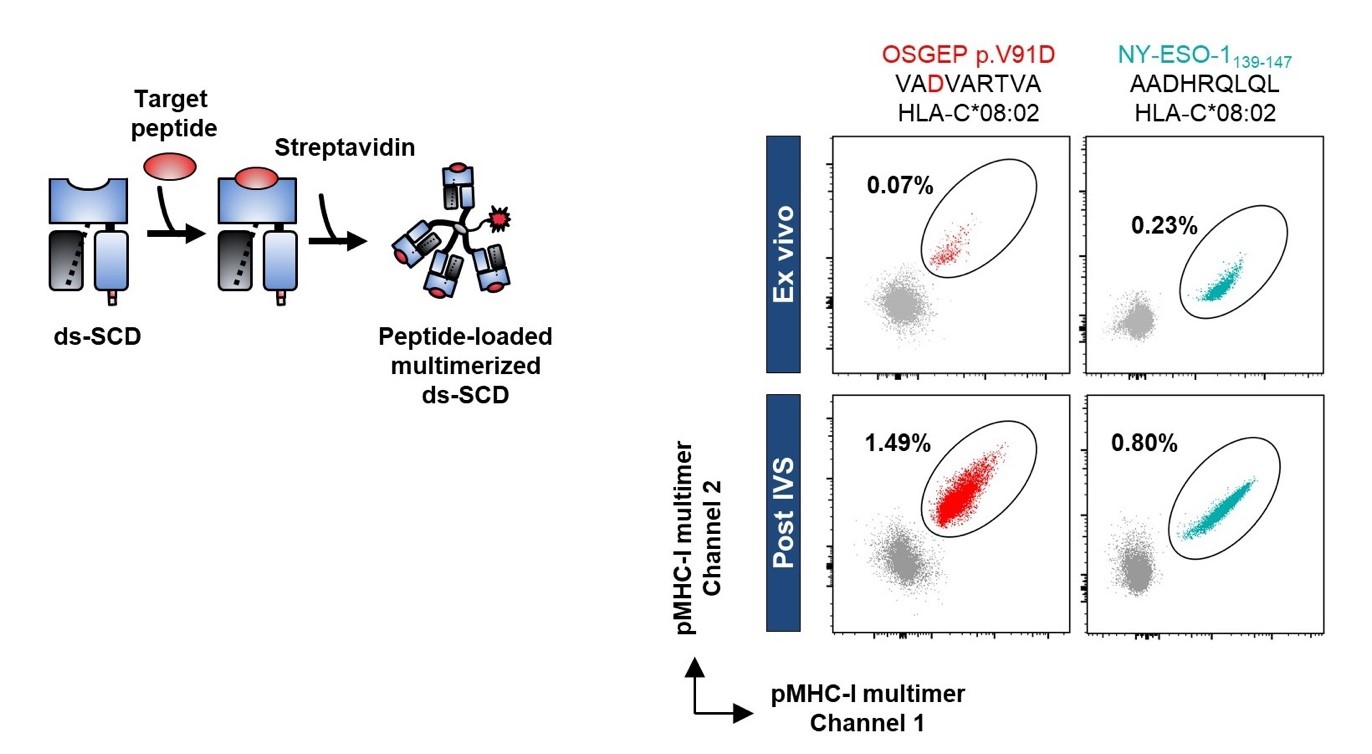
Left: Graphical presentation of disulfide-stabilized beta2-microglobulin HLA-A,B,C heavy chain fusion protein (ds-SCD). Ds-SCD can be loaded with peptides fitting with the used HLA-A,B,C allotype and multimerized with fluorophore-tagged streptavidin. Right: Identification of tumor peptide-reactive T cells in the peripheral blood from a melanoma patient. HLA-C*08:02 ds-SCD were loaded with either a neo-epitope peptide bearing a tumor-specific point mutation (left) or a peptide derived from the cancer-testis antigen NY-ESO-1 (right). pMHC-I multimer-labeled CD8+ T cell populations were detected ex vivo and expanded after a peptide pulsed-based in vitro stimulation (IVS).
© DKFZ
Team Members
Frank Momburg, Priv.-Doz. Dr. med. (Group head)
f.momburg@dkfz-heidelberg.de
Marten Meyer, Dr. rer. nat.
marten.meyer@dkfz-heidelberg.de
Márcia Figueiredo Gonçalves, Dr. sc. hum.
m.goncalves@dkfz-heidelberg.de
Annkathrin Teschner, Dr. rer. nat.
a.teschner@dkfz-heidelberg.de
Karsten Warwas, Dr. med.
k.warwas@dkfz-heidelberg.de
Christina Parpoulas, M.Sc. (former member)
Titouan Barthélémy, B.Sc.
titouan.bartelemy@dkfz-heidelberg.de
Nadja Bulbuc, technician
n.bulbuc@dkfz-heidelberg.de
Selina Börsig, technician
selina.boersig@dkfz-heidelberg.de
Claudia Tessmer, technician
c.tessmer@dkfz-heidelberg.de