Overview
These two layers of cell growth are interconnected, with signaling pathways controlling metabolism, and metabolites controlling signaling.
The Teleman Lab is interested in understanding cell and tissue growth. Compared to the cell cycle, the mechanisms controlling cell growth and tissue growth are less well understood.
We study two aspects of this problem: metabolism and signaling.
Metabolism: To grow, cells and tissues need to make biosynthetic building blocks to accumulate mass. This requires the activation of metabolic pathways that support anabolism and fight oxidative stress.
Signaling: Cellular anabolic metabolism is regulated by signaling pathways such as the insulin pathway or the mTORC1 pathway. These signaling pathways sense and respond to growth factors and nutrient levels. We investigate the regulation of these signaling processes controlling metabolism and energy consumption.
This research has implications for both basic biology and cancer biology:
Basic biology: Tissue growth and size control represents one of the unsolved fundamental mysteries in developmental biology. How does a tissue know what size to become, and when to stop growing? How does a tissue measure its size? The underlying mechanisms are not known.
Cancer biology. To form a tumor, cells need to both proliferate and grow. Understanding the molecular mechanism controlling cell growth, and the metabolic pathways that generate biomass, is an important part of understanding cancer.
Research on tissue and cell growth covers many different topics. As detailed below, some of the topics we study are the functional characterization of new cancer genes, novel translational control mechanisms, or the regulation of signaling pathways by metabolites. Since signaling pathways are highly conserved from flies to human, we use Drosophila as a model organism for gene discovery and for organismal studies. We then translate our findings into mammalian systems to study the implications for metabolic disorders and cancer. We do this in human cell culture and in clinical studies with the Heidelberg University Hospital.
Discovering the function of cancer genes
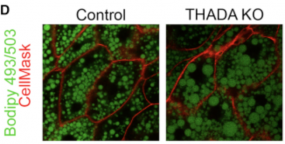
Fat body of THADA KO animals stained for neutral lipids with Bodipy 493 reveals that they have more lipid per cell and larger lipid droplets compared to controls.
© dkfz.de
About 50% of all protein coding genes are of unknown function. While cancer genome sequencing discovers many mutations in novel genes, the functions of most of these genes are still unknown. Since the molecular mechanisms of cell growth, metabolic regulation, transcriptional regulation and protein translation are conserved across vast phylogenetic distances, we use the model organism Drosophila to uncover new gene functions in health and disease. Indeed, much of our understanding of signaling pathways and their components comes from Drosophila research.
For example, with Drosophila knock-outs we were able to discover the function of the cancer related gene THADA. THADA was a gene found mutated in thyroid adenomas, and genome-wide association studies (GWAS) identified THADA as one of the top genes that was evolutionarily selected in response to cold adaptation in human populations, as well as one of the top genes associated with type 2 diabetes. Furthermore, sequencing of the Neaderthal genome identified THADA as one of the genes most strongly selected in modern humans. The molecular function of THADA, however, was not known.
We found that THADA regulates the balance between body heat production versus energy storage, and found that it does so via the ER calcium pump SERCA. When THADA activity is low, animals become fat, and dont produce enough heat so they are cold sensitive.
This provides some of the first evidence that obesity may be associated with an evolution in humans towards reduced heat production. Indeed, predisposition for obesity is more wide-spread in human populations that have adapted to warmer climates, leading to the hypothesis that in warm climates the human body reduces its metabolic rate to reduce heat production.
Furthermore, this discovery indicates that THADA controls cell metabolism and energy consumption. Hence it will be interesting to understand in the future how these metabolic alterations explain the involvement of THADA in cancer development.
Cancer sequencing is identifying mutations in many novel cancer genes of unknown function. It is an exciting time to use Drosophila genetics to discover what these cancer genes are doing functionally.
Reference
Moraru A, Cakan-Akdogan G, Strassburger K, Males M, Mueller S, Jabs M, Muelleder M, Frejno M, Braeckman BP, Ralser M, and Teleman AA. THADA regulates the organismal balance between energy storage and heat production. (2017) Developmental Cell 41: 72-81 [pubmed]
Funding
This work was funded by the Deutsche Krebshilfe and the German Research Foundation SFB1118.
How cells and organisms sense metabolites
Regulation of mitochondrial morphology and function by stearic acid C18:0
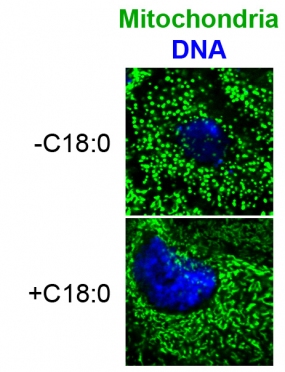
The images show larvae cells stained with mito GFP (green) and DAPI (blue). Starvation of larvae causes mitochondrial fragmentation, rescued by dietary C18:0.
© dkfz.de
Mitochondria are not only the powerhouse of the cell, but they are also involved in a variety of cellular functions, including amino acid and lipid biogenesis and breakdown, signaling and apoptosis. Mitochondrial dysfunction has been linked to neurodegenerative disease, cancer and aging. Little is known how mitochondrial function is regulated.
We identified the metabolite C18:0 as a signaling molecule regulating mitochondrial function and morphology in response to diet in Drosophila and human cells. We discovered that C18:0 is covalently attached to a cell surface receptor which signals via a new pathway to control mitochondrial fusion and function.
This work was done in collaboration with the lab of Antonio Vidal-Puig at the University of Cambridge and the lab of Aubrey Miller at DKFZ.
We have translated our findings into humans via a clinical study in collaboration with the Nawroth lab at Heidelberg University Hospital. The study shows that our bodies are sensitive to the amount of stearic acid (C18:0) in our diets. The amount of stearic acid that we eat affects mitochondrial morphology and function in our cells within a few hours after eating it, thereby affecting our physiology. This has implications for human health and diet.
Selected references
Senyilmaz-Tiebe D, Pfaff DH, Virtue S, Schwarz KV, Fleming T, Altamura S, Muckenthaler MU, Okun JG, Vidal-Puig A, Nawroth P, Teleman AA. Dietary stearic acid regulates mitochondria in vivo in humans. (2018) Nature Communications 9:3129. [article]
Protein stearoylation
We just received an ERC Consolidator Grant to study protein stearoylation.
One way metabolites can affect signaling pathways is by forming direct covalent modification on proteins. For instance, protein acetylation involves the attachment of an acetyl group from Acetyl-CoA onto a protein. Likewise, we found that C18:0 covalently binds to the cell-surface receptor Transferrin Receptor TfR1 leading to a signaling cascade that targets mitochondrial fusion and thereby function, both in Drosophila and in human cells. To our knowledge, this is the first time stearoylation of a human protein was shown to regulate its function.
We are working to characterize the stearoylome in Drosophila and humans, identifying which proteins are stearoylated by C18:0, and how this affects cellular signaling and physiology. This work will link our diet to responses in cell signaling and organismal physiology.
Selected references
Senyilmaz D, Virtue S, Xu X, Tan CY, Griffin JL, Miller AK, Vidal-Puig A and Teleman AA. Regulation of mitochondrial morphology and function by Stearoylation of TfR1. (2015) Nature 525:124-8 [pubmed]
Dynamic regulation of TOR by amino acids
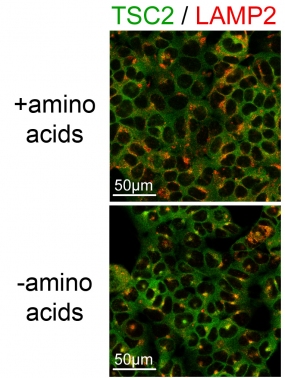
The images show the TSC2 localization in cells by immunostaining of TSC2 (green) and the lysosomal marker LAMP2 (red). Amino acid starvation (-amino acids) causes lysosomal re-localization of TSC2 (yellow).
© dkfz.de
mTOR is a central regulator of cell growth. Highly conserved from yeast to humans, it is one of the most powerful anabolic signals in a cell. When mTOR is active, cells accumulate mass and grow, whereas when TOR is inactive, cells cannot grow.
For this reason, mTOR is dysregulated in almost all human cancers. mTOR promotes cell growth by inducing protein synthesis, as well as synthesis of other cellular building blocks such as lipids for the cell membrane. Because of its central role in regulating growth, mTOR activity is regulated by a large number of inputs including growth factor signaling (such as insulin), cellular stresses (such as hypoxia, or high AMP levels) as well as the availability of amino acids.
The molecular mechanisms by which some of these inputs regulate mTOR, however, are still not understood. We could show that both amino acids and growth factors are required to avoid mTORC1 inhibition by the TSC1/2 complex. The presence of any single stress is sufficient to cause TSC2 lysosomal localization and inhibition of TORC1. This effect is not cell-type specific, indicating an underlying general biological detection mechanism to a variety of stress stimuli.
A few projects in the lab aim at identifying novel regulators of TOR, in particular relating to cellular amino acid sensing, and to understand their function both at the molecular and the physiological level.
Selected references
Demetriades C, Doumpas N, and Teleman AA. Regulation of TORC1 in Response to Amino Acid Starvation via Lysosomal Recruitment of TSC2. (2014) Cell. 156:786-99 [pubmed]
Plescher M, Teleman AA and Demetriades C. TSC2 mediates hyperosmotic stress-induced inactivation of mTORC1. (2015) Scientific Reports 5:13828 [pubmed]
Tsokanos F-F, Albert M-A, Demetriades C, Spirohn K, Boutros M, and Teleman AA. eIF4A inactivates TORC1 in response to amino acid starvation. (2016) EMBO J. 2016 May 17;35(10):1058-76 [pubmed]
Demetriades C, Plescher M, and Teleman AA. Lysosomal recruitment of TSC2 is a universal response to cellular stress. (2016) Nature Communications 7:10662. [pubmed]
Funding
These projects are part of the local Collaborative Research Center SFB1036.
Cancer and non-canonical translation

DENR-KO die as pharate adults with larval-like abdominal epidermis. DENR promotes cell proliferation and boosts protein synthesis in proliferating cells.
© dkfz.de
To grow, cancer cells need to synthesize proteins. Cancer cells, however, face a challenge. Canonical textbook cap-dependent translation is turned off in response to cellular stresses. Hypoxia, nutrient limitation, ER stress, genotoxic stress and cytotoxic stress all inhibit canonical translation (by phosphorylating eIF2alpha and by inhibiting mTORC1, which leads to 4E-BP activation and eIF4E inhibition). Since almost all cancer cells have one stress or the other, they need to either bypass these inhibitory mechanisms, or to rely on alternate non-canonical translation mechanisms. We are studying some of these non-canonical translation mechanisms.
For instance, we recently discovered that the oncogenes DENR and MCT-1 promote a process called translation re-initiation, to promote expression of genes required for proliferation. Several projects in the lab focus on how translation is regulated, and how its dysregulation contributes to cancer.
Selected references
Ahmed YL, Schleich S, Bohlen J, Mandel N, Simon B, Sinning I, Teleman AA. DENR-MCTS1 heterodimerization and tRNA recruitement are required for translation reinitiaion. (2018) PLoS Biol16(6):e2005160. [pubmed]
Schleich S, Strassburger K, Janiesch PC, Koledachkina T, Miller KK, Haneke K, Cheng Y-C, Kuechler K, Stoecklin G, Duncan KE and Teleman AA. DENRMCT-1 Promotes Translation Reinitiation Downstream of uORFs to Control Tissue Growth. (2014) Nature 512: 208-212 [pubmed]
Schleich S, Acevedo JM, Clemm von Hohenberg K and Teleman AA. Identification of transcripts with short stuORFs as targets for DENRMCTS1-dependent translation in human cells. (2017) Scientific Reports 7:3722 [pubmed]
Reactive carbonyl species and causes of diabetes
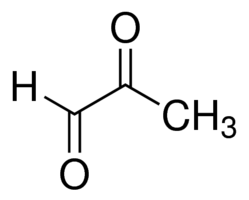
Methylglyoxal (MG)
One set of phenotypes associated with type 2 diabetes is insulin resistance and hyperglycemia. In fact, insulin resistance is often considered to be the cause of type 2 diabetes. Indeed, Type 2 diabetes is treated with insulin sensitizers or with insulin. Unexpectedly, however, recent clinical studies find that therapies renormalizing blood glucose to non-diabetic levels does not prevent progression of the disease patients still develop neuropathies, nephropathy and retinopathy. This suggests that insulin resistance is one phenotype of type 2 diabetes, but not the cause. This raises the important question: what are the causes of type 2 diabetes?
We are working on this question in collaboration with the Nawroth lab at Heidelberg University Hospital. Data from our lab suggest one possible cause could be elevated levels of the metabolite methylglyoxal. Methylglyoxal, which forms as a byproduct of several metabolic pathways, forms adducts on proteins to alter their function. We are studying the detoxification of methylglyoxal, and the consequences of impaired detoxification for diabetes.
Selected reference
Moraru A, Wiederstein J, Pfaff D, Fleming T, Miller AK, Nawroth P and Teleman AA. Elevated levels of the reactive metabolite methylglyoxal recapitulate progression of Type 2 Diabetes. (2018) Cell Metabolism doi: 10.1016/j.cmet.2018.02.003 [pubmed]
Pfaff DH, Fleming T, Nawroth P and Teleman AA. Evidence against a role for the Parkinsonism-associated protein DJ-1 in methylglyoxal detoxification. (2016) JBC 292:685-690 [pubmed]
Funding
These projects are part of the Collaborative Research Center SFB1118.
Understanding growth control during development
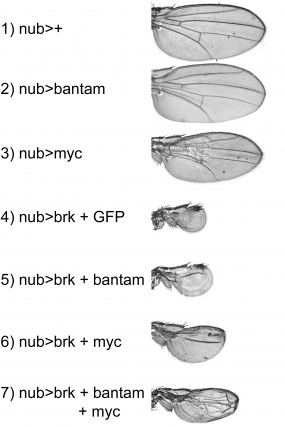
Wings expressing Brk ubiquitously during development are strongly reduced in size, and lose patterning elements such as veins. Reconstitution of bantam or Myc expression either alone or in combination rescues the undergrowth caused by Brk, but not the patterning defects.
© dkfz.de
During animal development tissues increase tremendously in mass, growing roughly 1000-fold from the embryo to the adult. Each tissue then stops at a very precise size, which is determined by a combination of genetic and environmental factors. The regulation of this process is a fundamental open question in developmental biology. Furthermore, it is of relevance since the molecular pathways that control growth are linked to the re-initiation of cell growth and proliferation observed in cancer.
In this context, we are studying signaling pathways that regulate tissue growth. These include: (1) Insulin / PI3K / mTOR (2) Hippo, Yorkie/YAP (3) and Dpp (TGF-beta in mammals).
One model system we work with is the fly wing, since it is a well-studied model system for tissue growth. The wing is specified as a group of 50 cells which then proliferate to yield a tissue with roughly 50,000 cells. When it reaches the correct final size, the cells in the wing stop proliferating. The underlying molecular mechanisms are not known. We are interested, for instance, in understanding how the Dpp morphogen gradient controls growth. Furthermore, we aim to discover novel genes and interactions regulating this process.
We could show that TORC1 activity in the Drosophila wing disc is activated by the Dpp pathway, a signaling pathway that strongly affects tissue size, and that TORC1 activity is tied into the cell cycle via CycD/Cdk4.
Selected references
Romero-Pozuelo J, Demetriades C, Schroeder P and Teleman AA. CycD/Cdk4 and discontinuities in Dpp signaling activate TORC1 in the Drosophila wing disc. (2017) Developmental Cell 42:376-87 [pubmed]
Doumpas N, Ruiz-Romero M, Blanco E, Edgar B, Corominas M and Teleman AA. Brinker regulates wing disc growth in part via repression of Myc expression. (2013) EMBO Reports. 14:261-8 [pubmed]
Teleman AA, Cohen SM, Dpp gradient formation in the Drosophila wing imaginal disc. (2000) Cell. 2000 Dec 8;103(6):971-80 [pubmed]
Aging
Why do we age? Can we extend the period of our lives during which we are healthy our healthspan?
Metabolic homeostasis has been linked to aging in multiple ways. For instance, dietary restriction is one of the main lifestyle interventions known to delay aging. Exercise also delays aging and this occurs via modulation of cellular metabolism as well as signaling through nutrient-responsive pathways such as AMPK. Thirdly, nutrient supply regulates acetyl-CoA levels in cells, thereby affecting histone acetylation, which has been linked to aging. Finally, one of the main nutrient-sensitive kinases, mTORC1, is a potent modulator of aging in multiple animals including flies and mice. Indeed, a recent clinical study by Novartis showed that mTORC1 inhibition can partially revert one aspect of aging in humans - immunosenescence.
Since we have a focus on metabolic homeostasis, the biological processes that we study impact aging: mTOR signaling, mitochondrial function, and detoxification of reactive carbonyl species. Several projects in the lab analyze the impact of these processes on animal aging.
Funding
The Helmholtz Vernetzungsfond has allocated funding for the project Aging and Metabolic Programming which is coordinated by the lab head A. Teleman and S. Herzig, director of Institute for Diabetes and Cancer (IDC) at HMGU in Munich. The project´s aim is to establish a nationwide research network across the Helmholtz Health Centers to combine expertise in various diseases to explore age-related metabolic dysfunction and the development of therapeutic measures. Please visit the AMPro website for more information.